私たちは空間における体の各部位の位置を意識的に把握しているのは明らかですが、脳が位置情報を計算するための入力となる筋肉や関節からの信号は、私たちの意識的な認識には届いていないと考えられることが多いのです。肌への触覚は感じることができます——それは確かです。しかし主流の神経科学によると、痛みや緊張を除いて、私たちは筋肉に「触覚的」な感覚を感じることはできないようなのです。しかし、本当にそうなのでしょうか?筋肉に、ニュートラルあるいは心地よい感覚を感じたことが、本当に一度もないのでしょうか?もし正直な答えが「いいえ、そんな感覚はないと思います」であったとしても——それもまた興味深い回答で、この記事はあなたにも向けられています。この記事では、身体認識の一次元としての「固有受容感覚的意識」、その潜在的な神経基盤、そしてなぜ——実在し、訓練によって発達できる知覚能力であるにもかかわらず——科学文献に確立された名称が存在しないのかについて掘り下げていきます。
活動中の筋肉に宿る局所感覚——痛みでもなく、筋肉痛でもなく、ただ筋肉が働いている感覚——には、科学文献に確立された名称が存在しない。
はじめに
もし身体運動の利点を重要度の高い順に挙げるよう求められたとしたら、知覚能力の発達——筋肉の感覚と、体の表面および内部にある複数の点の位置を常に意識できる能力——を最初に挙げるでしょう。体調を整える、健康になる、気分が良くなる、生産性が上がる、といった他のあらゆる利点は、これに自然に続いてくるように思えるのです。
しかし、私の考える第一位の利点が、こうしたリストに載ることはほとんどありません。運動があなたにもたらすものを列挙したカタログは、知覚的な効果より先に健康上の効果を挙げます——そもそも知覚的な効果に触れるとしたら、の話ですが。
広く認識されている「身体意識」というカテゴリは、私の第一位と重複する部分があります。「身体意識の利点」については、かなりの文献が存在します。しかしこの用語は一貫性なく使われており、明確な定義が示されることはほとんどないのです。運動感覚から、Craigの内受容感覚、さらにはEllen Langerのマインドフルネスに近いものまで、様々なものを指す場合があるのです。
私の第一位は、これらのいずれでもありません。私たちの枠組みでは、それは「固有受容感覚的意識」と「空間意識」の組み合わせを指しています。この記事では固有受容感覚的意識のみに焦点を当てます。「身体意識」の共通定義がまだ存在しないため、まずはこの概念について語るBaseworks独自の枠組みを紹介したいと思います。
身体認識は、より微妙な概念化を必要とする複雑な多次元構造です。明確な定義が提供されることは滅多にありません。
3種類の身体意識
Baseworksでは、実用的・教育的観点から、身体認識の3つの「種類」または「次元」を区別することが有益です。
- 内受容感覚意識
- 固有受容感覚的意識
- 空間的身体意識
内受容感覚的意識とは、内臓の機能、ストレス、感情、代謝の変化に関連する感覚の意識的な認識のことです。私たちの内受容感覚的意識の概念は、Price & HoovenによるCraigの内受容感覚の考えを知覚スキルとして拡張したものと、ほぼ一致しています。
残りの2種類はより独特であり、既存の枠組みや分類と正確に一致させることは困難です。
Baseworksの枠組みにおける空間的身体意識とは、最も基本的な次元では、視覚入力なしに体の各部位の位置を感じとる能力(運動感覚的局在化)を指します。これに加え、Baseworksの枠組みでは、空間意識は空間のメンタルモデルを外部環境にマッピングし、様々な目的——記憶を助ける、美的、探索的、レクリエーションなど——に活用する能力とも関連しています。これは私の第一位に関連していますが、この記事では取り上げません。
Baseworksの枠組みにおける固有受容感覚的意識とは、(皮膚ではなく)筋骨格系の機械受容器からの信号の意識的な認識であり、局所化された感覚として経験されるものです。
注:皮膚の触覚感覚は「外受容感覚」(外部環境を感知すること)の領域に属し、3種類の身体意識の枠組みには含まれません。
この分類は、応用・タスク志向のものであり、特定の感覚をどう分類すべきか不明確な場合があります。
たとえば、心拍を意識しているとき——指先で皮膚に触れることなく、体のどこかに脈動する感覚がある場合——これは内受容感覚的意識なのか、それとも外受容感覚的意識なのでしょうか?大きな血管には特殊な伸展感受性受容体が備わっていますが、Wilfrid Jänig(自律神経系に関する私の最も頼りになる情報源)によると、それらの刺激は意識的な感覚を引き起こさないようなのです。したがって、心拍の感覚は実際には皮膚によって捉えられているのかもしれません(つまり、外受容感覚的能力が内受容感覚的意識に寄与していると言えるかもしれません)。
あるいは、横行結腸の蠕動運動——おへそのすぐ上にある大腸のゆっくりとしたリズミカルな収縮——が腹筋の内側で動くのを意識した場合(偶然かどうかは不明ですが、東京で鍼灸師を訪ねるたびに私が最初に針を打たれる場所です)、これは内受容感覚的意識と見なすべきか、それとも固有受容感覚的意識と見なすべきなのでしょうか?
明確な答えがない場合もありますが、この枠組みは指導応用と研究のニーズに応えるものです。また、「身体意識」について議論する際に生じがちな2つの問題を解決するのにも役立ちます。
- 無関係な現象を混ぜ合わせる傾向(例:「身体意識とは空間における体の位置への意識であり、空腹への意識も含まれる」)
- 既存のカテゴリの境界線上に曖昧に位置するために特定の現象に気づかない傾向(例:「固有受容感覚的意識は意識の及ばないところで発生する」)
固有受容感覚と固有受容感覚的意識
本題に深く入る前に、重要な用語の区別に注目しておきたいと思います。
固有受容感覚と固有受容感覚的意識は、頻繁に同義語として使われているにもかかわらず、同じものではありません。
固有受容感覚とはシステムそのものです——筋肉の長さ、力、関節の位置に関する情報を神経系に継続的に供給する受容器、神経路、そして脳の回路のことです。その多くは私たちが気づかないうちに起きており、なくてはならないものです。固有受容感覚がなければ、動作を協調させること、バランスを保つこと、目で見ずにグラスに手を伸ばすことさえできないでしょう。
固有受容感覚的意識は、私たちがこの言葉を使うとき、異なるものを指します。それは、このシステムによって生み出される感覚への意識的なアクセスであり、筋肉や関節の中に局在化できる感覚として経験されるものです。
知覚システムとその信号への意識的な認識との間のこの区別は、新しいものではありません。内受容感覚の文献でも同様の区別がなされています。心拍数、腸の蠕動、内臓の状態といった深い内受容感覚的信号は、私たちの意識を介さずに作動しますが、内受容感覚的意識——それらの信号に意識的に注意を向け、表現する能力——は訓練可能なスキルであることが示されています(Price & Hooven, 2018)。以下で論じるように、固有受容感覚についても同様のことが言えるかもしれないのです。
固有受容感覚的意識、本当に?
「固有受容」という用語には、幅広い意味があります。例えば神経科学者は、身体的固有受容を完全に非個人的な、無意識の機能として扱う場合があります。
— Shaun Gallagher (身体化された認知、社会的認知、主体性、精神病理学の哲学に関する研究で知られる哲学者)
意識的な感覚として定義される「固有受容感覚的意識」に眉をひそめる方は少なくないでしょう。固有受容感覚システムは、意識的に局在化できる感覚を生み出せるのでしょうか?
指が痛い場合や誰かが指に触れた場合、その感覚を首や胃ではなく指に局在化することができます。それが私の言う「局在化できる」という意味であり、これは広く共有された経験です。しかし教科書には、固有受容感覚はそのように機能しないと書かれています——意識的に局在化できる感覚の閾値以下で作動するのだ、と。
「身体認識」に関する科学文献の相当な部分は、主に内受容感覚的意識に焦点を当ててきました。2009年の身体意識評価ツールに関するシステマティックレビューでは、固有受容感覚的な次元が現れるのは、心理的苦痛の代理指標として——「苦悩、心配、痛み、緊張の感覚(例:「私は筋肉の緊張に気づいています」)」——だけであり、独自の領域としてのニュートラルあるいはポジティブな感覚としてではないことがわかりました(Mehling et al., 2009)。
それでも、
痛みや筋肉痛以外に、筋肉に何かを感じたことが本当にないのでしょうか?
同じグラスを2つ用意してください。一方を室温の水で満たしてください。両手に一つずつ持ち、目を閉じます。どちらが重いかはすぐにわかります。しかし、そうしている間に腕の感覚に注意を払えば、別の何かに気づくかもしれません——重いグラスを持っている腕に、何かがより多くある感覚。ぼんやりとして拡散しているが、筋肉の中に局在化できる感覚です。
これが、この記事が取り上げる「謎の」感覚です。
この時点で、読者の受け取り方が二つに分かれると予想しています。「この感覚の何が不思議なの?普通の人間の体験じゃないの?」と戸惑う方と、「一体何の話をしているの?」と、これも戸惑う方です。この両方の反応に答えていきたいと思います。
私たちが非実践者を対象に行っている継続的な調査(現在n=48)では、回答者の約47%が安静時に筋肉の感覚を意識していると報告し、約47%が運動中のみ意識すると報告し、残りの6%は身体活動中でも筋肉に局所感覚を感じないと報告しています。さらに、安静時の感覚を知っている回答者のほぼ半数——48%——は、この感覚を何と呼べばよいかわからないと回答しました。目の見える人が光や色を語る語彙を持っていないようなものです。

私たちの調査データと様々な人との対話から、この感覚を表すために最もよく使われる言葉は活性化であることがわかりました。これは筋肉の中に局在化した意識的な感覚であり、筋肉が収縮すると現れるか強まるものです。収縮している筋肉でのみ感じ、安静時には感じないという人もいます。即時的なもので——疲労とともに蓄積する感覚ではありません。痛みでも緊張でもなく、不快でもありません。何かをやめたくなるようなものではなく、使われている筋肉が持つ正常な質感のように感じられます。
科学文献には、私の知る限り、この感覚の名前がありません。
興味深いことに、哲学的な文献もこのギャップを埋めていません。メルロ=ポンティの伝統に立つ現象学者たちは、固有受容感覚を生きられた体験の次元として真剣に捉えてきました——空間を感じながら体を持つとはどのようなことか、方向感覚としての姿勢、意図的な身振りとしての運動、統一された存在としての身体、といった探求においてです。Gallagher自身の身体スキーマと身体イメージに関する研究もこの伝統に属します。しかしこれは固有受容感覚を異なる処理レベルで、異なる焦点から扱っています。その伝統が「固有受容感覚的現象学」と呼ぶものは、環境の中を動く際の空間的・運動感覚的体験を意味する傾向があります(これは私たちが空間意識と呼ぶ知覚領域に近いものです)——特定の筋肉の内側での感覚の質感ではなく。より具体的な問い——痛みやデッドリフト後の乳酸性疲労なしに、筋肉そのものはどのように感じられるのか——はそこではほとんど浮上しません。哲学的文献がそのような感覚は存在しないと主張しているわけではありません。ただ、異なる探求の角度を持っており、その問いはその中から生じてこないのです。
この記事の主な目的は、筋骨格系(主に筋肉)に起因する意識的な感覚に注目し、なぜこの現象が——実在し、訓練によって発達できる知覚能力として現れているにもかかわらず——科学文献でほとんど注目されてこなかったのかを探ることにあります。
次の図は、この記事の内容を要約しています。内受容感覚、固有受容感覚、外受容感覚について私たちが通常考える非常に規則的なシステムの中の、ブラックホール、盲点、空白について。
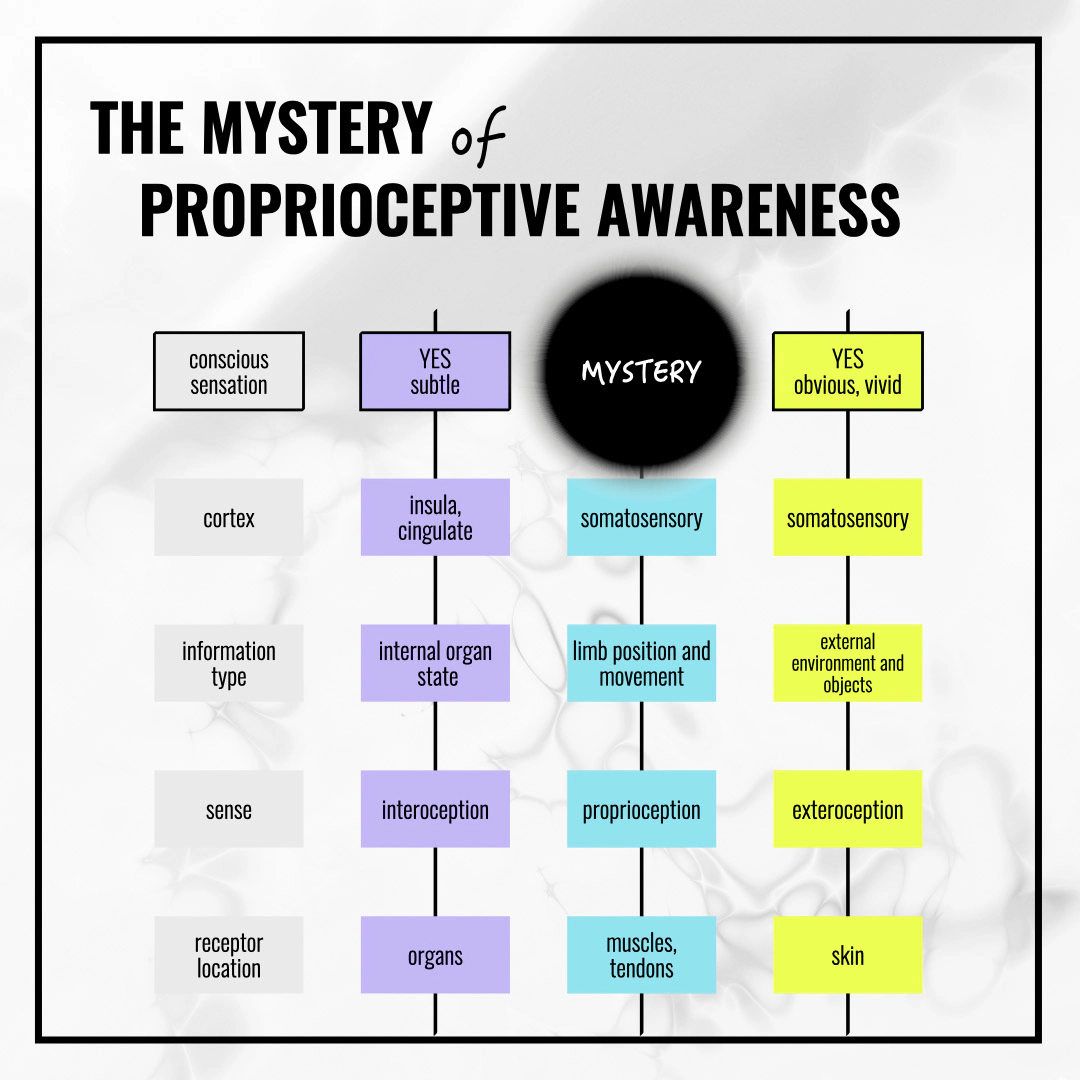
なぜそれは謎なのか?3つの二項対立
私たちの調査では、回答者のほぼ半数が、筋肉を能動的に収縮させなくても局所的な感覚を知覚できると報告しています。サンプルが、ウェブ検索を通じて身体意識関連のページにたどり着いた人々に偏っている可能性はありますが、それでもこの傾向は、これが広く見られる知覚的プロファイルであることを示しています。多くの人がこれらの感覚を知覚し、その描写に共感します。
しかし、「活動中の筋肉における意識的な感覚」——痛みでも筋肉痛でもなく、ニュートラルで局在化できる感覚——を科学文献の中で探しても、ほとんど何も見つからないのです。この話題が見落とされてしまう構造的な理由が、3つあると思います。
二項対立1:「意識的な」皮膚感覚と「無意識の」固有受容感覚
標準的な説明では、触覚——誰もが意識的で局在化可能なものと認めるもの——と、固有受容感覚——典型的には意識なく作動すると描写されるもの——を対比させます。この説明によれば、皮膚の受容器は脳に情報を送り、感じられる感覚を生み出します。筋肉の受容器は脳に情報を送り、運動指令と位置感覚を生み出しますが、感じられる感覚は生み出さない、ということになります。こうして、外受容感覚と固有受容感覚の間に二項対立が生まれます。
この見方は間違っているわけではありません。ただ不完全なのです。固有受容感覚の信号が脳によって無意識に位置座標に変換されるという事実は、筋骨格系からの信号が意識的体験に達することがないということを意味しません。しかし「皮膚」と「意識的感覚」を、「筋肉と関節の受容器」と「無意識の固有受容感覚」を一貫してペアとして扱うことで、標準的な枠組みは、痛みでも筋肉痛でもない筋肉の感覚のための概念的な場所を残さないのです。
この見解の好例として、このレビュー論文(Delhaye et al., 2018)が挙げられます。
二項対立2:「意識的な」固有受容感覚と「無意識の」固有受容感覚
固有受容感覚の中にも分裂があります。この説明によれば、後柱-内側レムニスカス(DCML)経路は、筋肉と関節からの体性感覚情報を皮質まで運び、意識的な位置感覚——目を使わずに腕がどこにあるかを感じとる能力——に寄与します。別の固有受容感覚の流れは小脳に入り、そこで信号が継続的なリアルタイムの運動協調のために使用されますが、伴われる意識はありません。
つまり、「意識的な」固有受容感覚と「無意識の」固有受容感覚について語るとき、皮質経路と小脳(皮質下)経路の間の区別が引かれることがあります。小脳処理は本当に無意識と見なされますが、位置感覚に入力するDCML経路は固有受容感覚の「意識的な」側面として特定されます。これもまた間違いではありませんが、局在化できる筋肉の感覚の余地を残しません。
この見解の好例としては、最近の書籍の章、特に図2(Vega and Cobo, 2021)や、TeachMeAnatomyの「The Ascending Tracts」ページなど、多くの一般向けの教育資料が挙げられます。
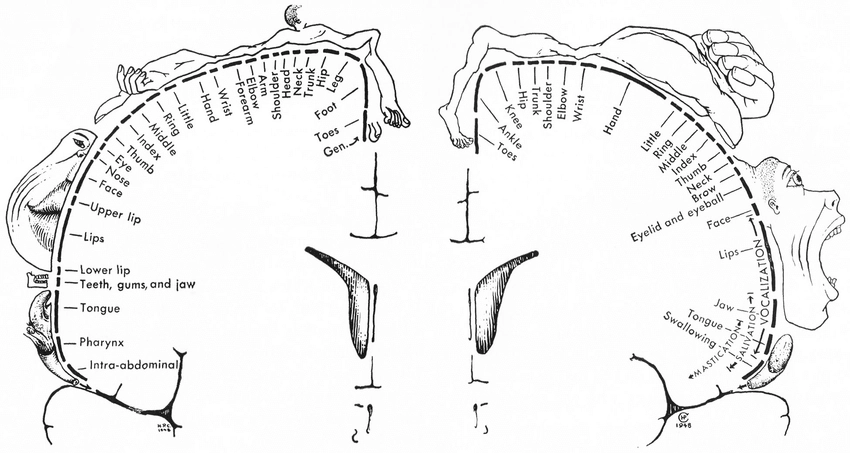
二項対立3:感覚-運動ホムンクルスの分裂
身体意識の話題に興味を持つ多くの人が、カナダの神経外科医Wilder Penfieldが、覚醒下開頭手術中にてんかん患者の脳を電極で刺激することで発見した感覚マップと運動マップのグラフィック表現である感覚ホムンクルスと運動ホムンクルス——脳皮質の細長い一片の上に変形した人が横たわっているように見える絵——を目にしたことがあるでしょう。この説明はほぼ常に触覚を基準として使います:「指に触れられたとき、どこに触れられたかわかるのは、このマップのおかげです」と。隣接する運動ホムンクルスは筋肉を出力として表します——「運動」側として。
これにより強力な暗黙の枠組みが生まれます:感覚=入力=皮膚、運動=出力=筋肉。筋肉もまた感覚を生み出す可能性があり、原理的には意識的認識に到達できるかもしれないという可能性は明示的に否定されているわけではありませんが、この枠組みが入出力の区別を強化することで暗黙的に抑圧されているのです。
この見解について明確な出典は存在しません。なぜなら、その前提が明示されることはめったにないからだ。むしろ、その前提は、神経科学の教科書や教育資料において、このトピックが一般的にどのように扱われているかの中に暗黙のうちに織り込まれています。
これら3つの二項対立は互いに強化し合います。合わさることで、構造的な盲点が生まれます:いずれの枠組みも明示的にこれを否定しているわけではありませんが、集合的に、筋肉における意識的で局在化できる感覚のための場所を残さないのです。この感覚はどの既存カテゴリにも属しません——触覚でも、痛みでも、位置感覚でも、無意識の固有受容感覚でもない——そのため、研究され、記述され、名付けられる傾向がないのです。
筋肉に関連する感覚は脳のどこにあるのか?
これらの感覚に一度も注意を払ったことがない人は、合理的にこう問うかもしれません:「なぜ意識的で局在化できる筋肉の感覚があるはずなのか?」
答えは、感覚信号が体から脳へと伝わる仕組みから始まります。神経系は専用の神経路を使います——特定の受容器から特定の皮質領域へ、特定の種類の情報を運ぶ専用のケーブルです。ここで最も関連する経路は、後柱-内側レムニスカス(DCML)経路です。
DCML経路は、2つの異なる源からの信号を緊密に並行して運びます:皮膚の機械受容器(圧力、振動、質感に反応する)と、筋肉・関節の機械受容器(伸展、力、関節の位置に反応する)です。どちらも直接、一次体性感覚野へ到達します——受容器から皮質まで3シナプスです。皮膚も同じ。筋肉も同じ。
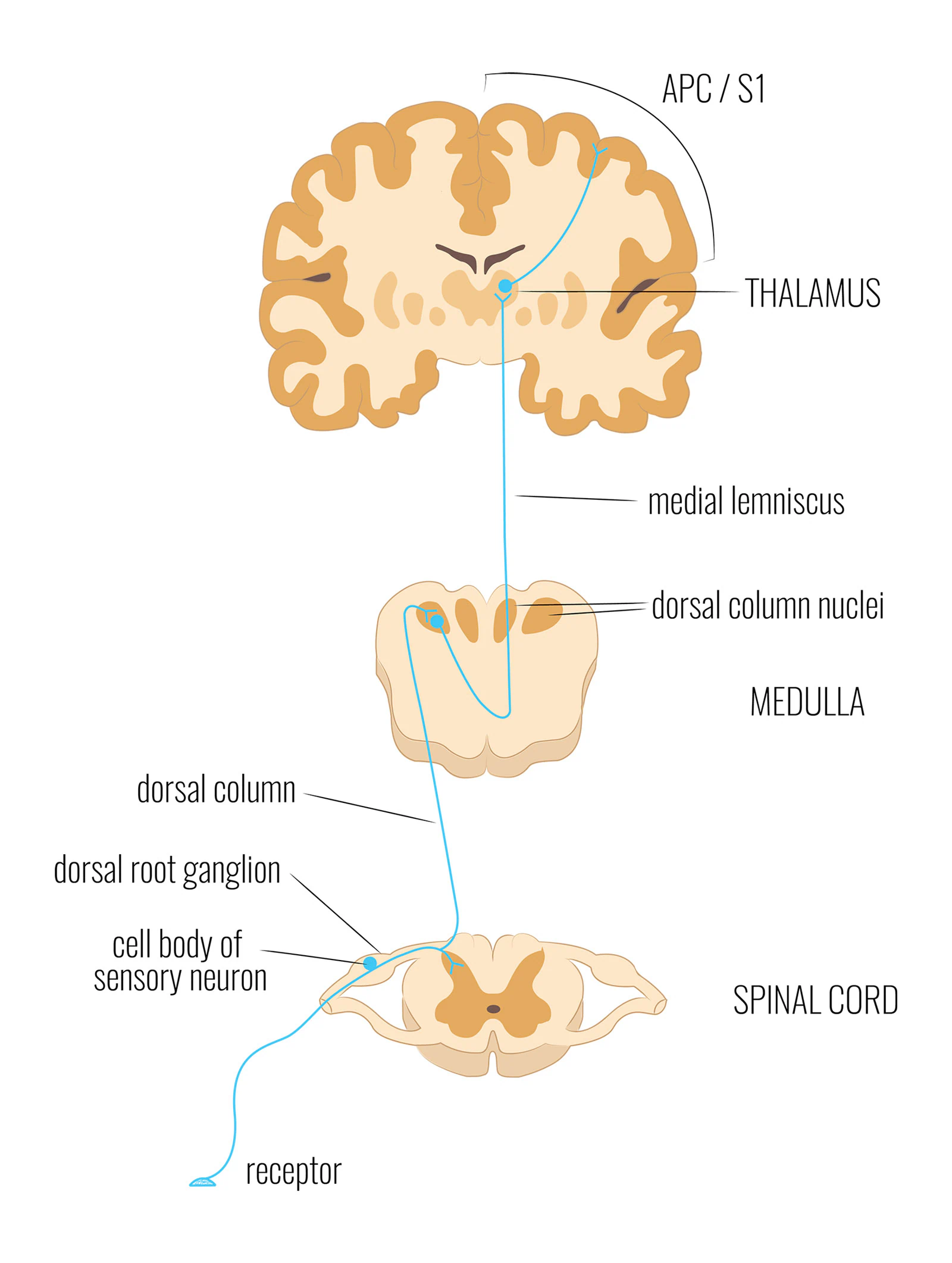
触覚については、これが意識的な局在化をどのように説明するかはよく知られています:皮膚からの信号は体の特定の位置に対応するS1の領域に到達し、体の特定の位置に触れられたという感覚を生み出します。S1には「皮膚表面のマップ」が含まれていると一般に言われています。
一次体性感覚野は単一の均一な領域ではありません。一般に「S1」と呼ばれるもの——より正確には前頭頂皮質——は4つの平行した帯で構成されています:3a野、3b野、1野、2野です。
- 3b野と1野は主に皮膚からの情報を受け取ります
- 3a野は主に固有受容感覚の入力を——筋肉と関節から——受け取ります
- 2野は皮膚と筋肉・関節の両方からの入力を受け取ります
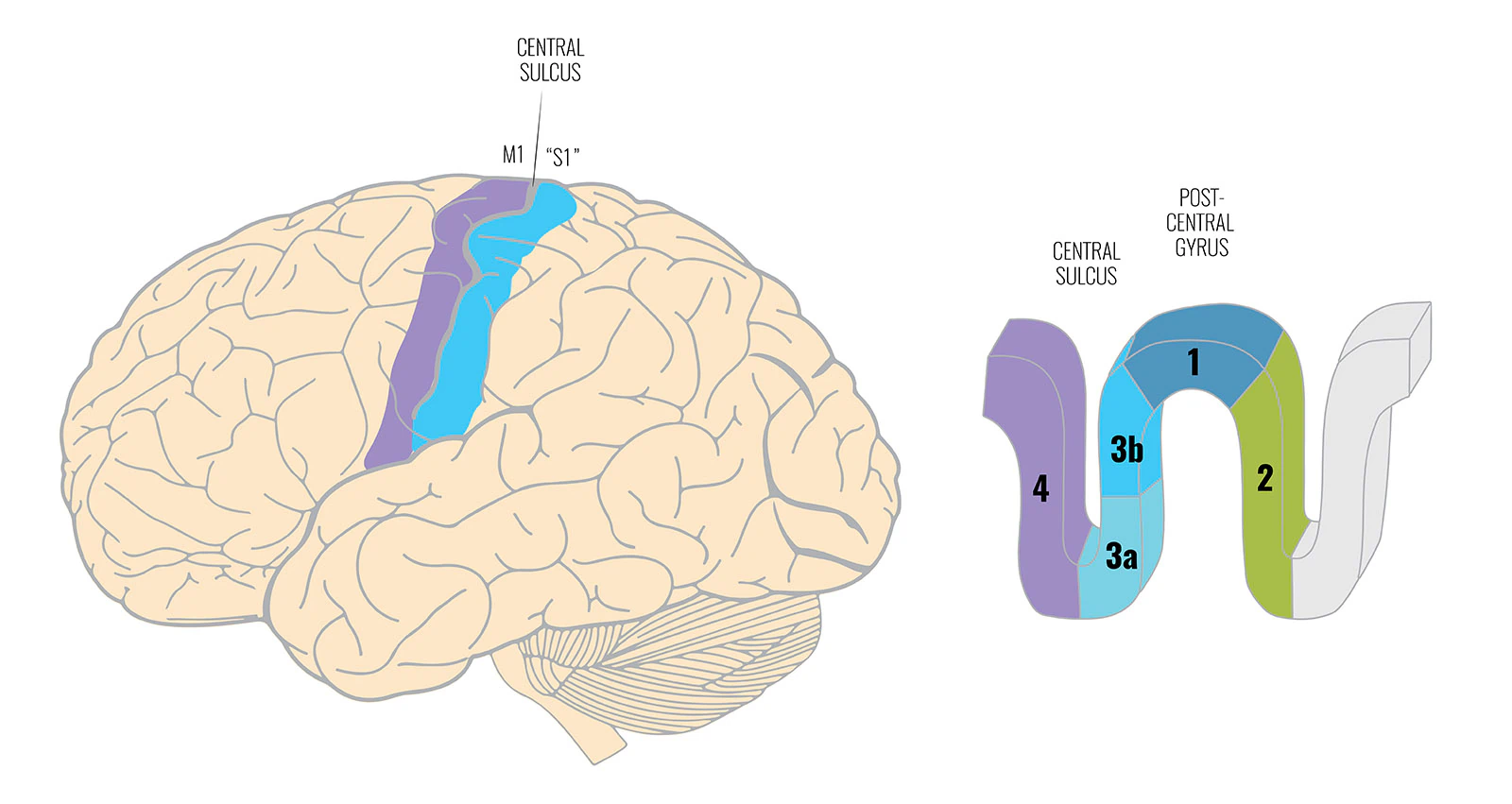
感覚ホムンクルスの標準的な説明は触覚を使います:誰かがあなたの手に触れたとき、このマップが信号を局在化するのでどこに触れられたかわかる、と。3b野と1野がその意識的な局在化の基盤です。すぐ隣の3a野が筋肉・関節の信号を受け取ります。
しかしここで、道は途絶えます。
皮膚の信号と筋肉の信号は同じ経路をたどり、同じ皮質の隣接領域に到達します。一方は明確に意識的で局在化できる感覚を生み出します。もう一方は、標準的な説明によれば、そうではない——何も感じることなく位置座標に変換される、ということになっています。健康で痛みのない人が3a野から実際に何を体験するのか——もし何かを体験するとしたら——という問いは、体系的に問われてこなかったようなのです。
固有受容感覚の第一線の研究者の一人が、個人的なやり取りの中でこのギャップを確認しました。彼自身は安静時の筋肉に感覚を経験しておらず、健康な被験者における持続的な意識的筋肉感覚のトピックに関する文献を知らないと述べました(個人的な連絡による)。そして付け加えました:「筋肉求心性線維の一次受容領域が3a野であることはあなたの言う通りですが、3a野を刺激すると安静時の筋肉に感覚が生じるという報告を読んだことはありません。」
3a野は、そのような感覚が処理される最も論理的な候補として残っています。しかし、この問いを決定的に解決するであろう実験は、この説明によれば、まだ行われていないようなのです。
この感覚とは何なのか?
では、この感覚は正確には何なのでしょうか?なぜ感じる人と感じない人がいるのでしょうか?この感覚にはどんな機能があるのでしょうか?気にかける必要はあるのでしょうか?感じていない人が見つけることはできるのでしょうか?
ここで述べることは、10年にわたる科学文献の調査、指導経験、そして様々な人との現象学的な対話に基づく仮説です。
意識的な感覚があるならば、受容器があるはずです。筋肉で最も豊富な受容器の種類は筋紡錘であり、しばしば伸展受容器と呼ばれます。筋紡錘は収縮を生み出す筋線維の間に位置し、筋肉が伸長すると発火率が上がります。この信号は脳に筋肉の長さと長さの変化速度を伝えるために送られ、脳はこの情報を使って関節角度と体の動き方を計算します。伸展受容器として、筋肉の伸長に反応し、一見すると、収縮とともに強まる感覚の候補としては不向きに思えます。
骨格筋にはまた、皮膚受容器に似た圧力受容器(パチーニ型受容器、ルフィニ型受容器)がいくつかありますが、はるかに少量しか存在しません。正直に言うと、数年前に初めてこれについて考え始めた時、私の最初の直感はこの感覚がこれらの圧力受容器に関連しているというものでした。
しかし、結論から言うと、筋紡錘の話は最初に見えるより複雑なのです。実際、筋紡錘を「受容器」として考えるのはやや誤解を招きます。生物学における受容器を考えるとき、特定のモダリティを信号として送る特殊化した細胞で、発火率は変わっても信号の種類は固定されているものとして概念化するかもしれません——眼の桿体と錐体のようなものです。しかし筋紡錘はそうではありません。筋紡錘は、複数の受容器を含む感覚器官として考える方が正確です。それは、本当に異なる仕事を持つ2種類の感覚終末を含むカプセルです:Ia群線維とII群線維で、それぞれ異なる種類の情報を脳に運びます。
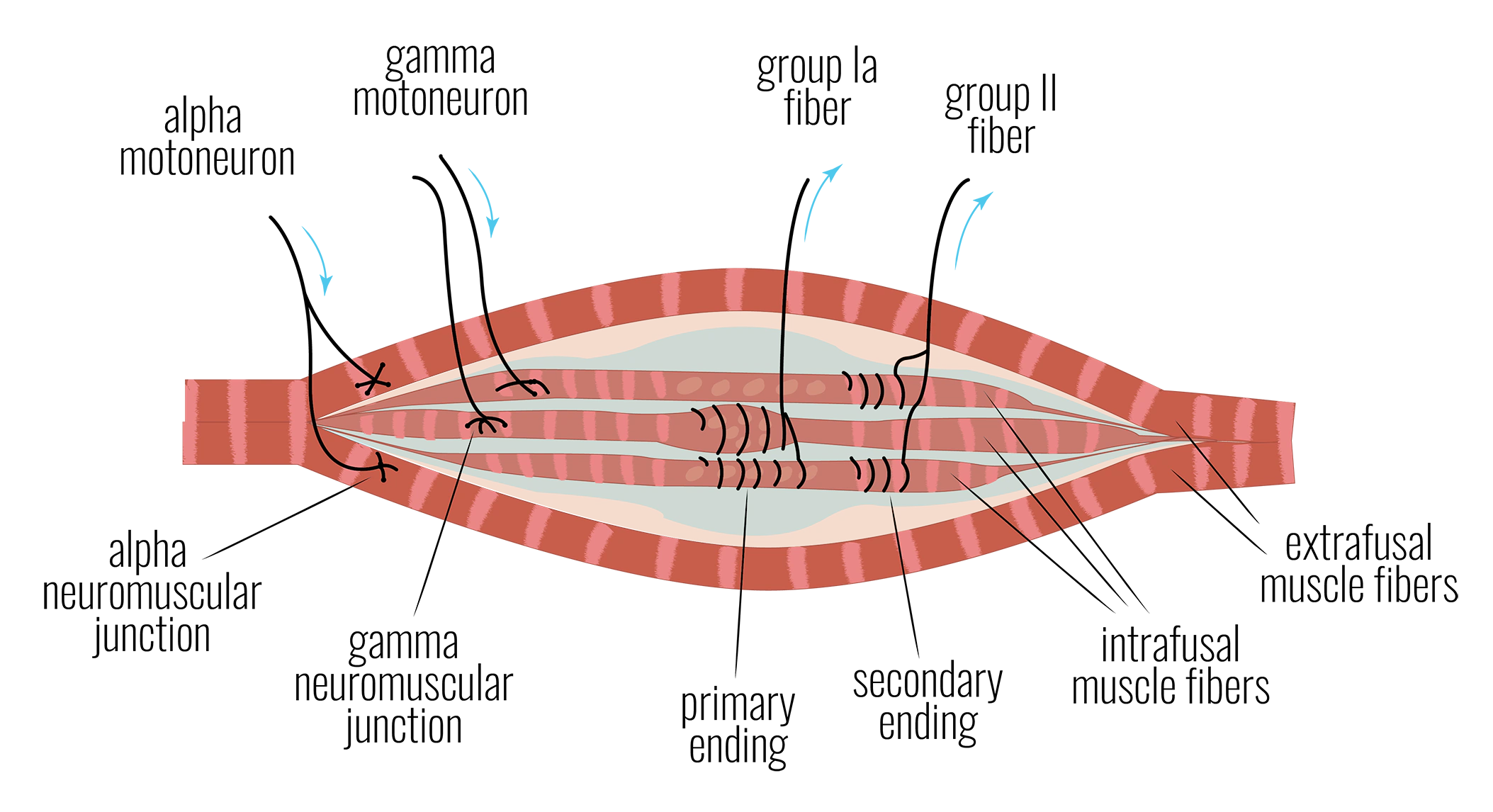
Ia群線維。 筋紡錘内の線維の中央部に付着しています。速度感受性があり、位相性です。筋肉が急に伸ばされると強く発火し、すぐに静まります。これが伸展反射の背後にある信号であり、筋紡錘が伸展受容器と呼ばれる主な理由です。
II群線維。 筋紡錘内の線維の末端部に付着しています。これは継続的に発火し、筋紡錘がどれほど張っているかに比例します——筋肉が能動的に伸ばされているか、しばらく静止しているかに関わらず(Vallbo, 1974)。この信号は安静時にゼロになりません。筋紡錘の現在の機械的状態を反映します。継続するために運動指令(進行中の動作)を必要としません。これが、筋肉における安静時のベースライン感覚のための最も妥当な基盤です。
なぜ脳は「筋紡錘がどれほど張っているか」を知る必要があるのでしょうか?ここがさらに興味深いところであり、筋紡錘の感覚の現象学がこれほど議論されていない理由でもあります。
筋肉が短縮すると、筋紡錘は通常たるんでしまいます——首を短くされたギターの弦のように。弦がたるんでいると音が出ません。たるんだ筋紡錘は信号を送れません。伸展(主な機能)に感受性を持つためには、筋紡錘は張っている必要があります。
神経筋系はこれを専用のメカニズムで解決しています。主要な収縮を生み出す筋線維に収縮を命令する運動ニューロン(α運動ニューロン)と並んで、別のニューロン群(γ運動ニューロン、錘内筋運動ニューロンとも呼ばれる)が筋紡錘の両端を収縮させ、周囲の筋肉が短縮する中でも中央部を張った状態に保ちます。α運動ニューロンとγ運動ニューロンは一緒に共活性化されます。これをアルファ-ガンマ共活性化と言います。その効果:張ったままの筋紡錘は発火を続けます——II群の緊張性信号とIa群の信号の両方が、収縮レベルに比例して強められます(高い力では感受性が低下することは後述します)。このメカニズムによって生成される求心性信号を、研究者たちは錘内筋再求心性入力と呼びます。(「求心性」とは「脳へと向かう信号」を意味し、「再求心性」とは、脳が生成した運動信号に応じて「脳へと戻る」信号を意味します。)
つまりII群の二次求心性線維は、「この筋紡錘が生きている筋肉がどれだけ伸ばされているか」ではなく、「この筋紡錘がどれほど張っているか」を脳に伝えます。これはある意味で維持管理的な信号であり、おそらくそれほど意識的な認識や注意を必要としないのかもしれません。考えてみてください:筋肉の筋紡錘がどれほど張っているかに基づいて決断を下した最後の機会はいつでしたか?その情報が必要なかったとしたら、単にそれに注意を払ったことがないという可能性が高いのです。
筋紡錘の図をGoogleで検索すると、α運動ニューロン(収縮を生み出す筋線維を収縮させる)とIa群求心性線維のみを示した図が多いことに気づくでしょう。これは筋紡錘の話がいつも語られる仕方を反映しています——筋肉が伸長したときに発火する伸展受容器として。γ運動線維は含むがII群線維は含まない図もあります——また「伸展受容器」の話として、つまり筋肉が短縮したときも「伸展受容器」として機能し続けるためにアルファ-ガンマ共活性化が必要だという文脈です。もちろん、II群求心性線維も適切に含む図もあります。それでも、インターネット上で表示されるすべての図に見られる傾向は、Ia群線維が主役を張る一方でII群求心性線維が二次的・補助的な役割を担うというものです。光を脳に運ぶ感覚線維を含め忘れた光受容器の図を見たことがないのです。つまり、II群線維の感覚の話は、支配的な「伸展受容器」の物語によって体系的に沈黙させられているのです。少し大げさに言っていますが、II群の「張り」信号が50年間測定されてきたにもかかわらず、その現象学は誰も問おうとしなかったということが、少し残念に思えるのです。
そして、これが、収縮中の筋肉におけるII群求心性線維の入力の知覚的相関に最も近づいた重要な論文につながります。
Luuと同僚たちが2011年に示したのは、錘内筋再求心性入力(筋肉が収縮するときに筋紡錘が脳に送る感覚信号)が、物体を持ち上げるときの力と重さの感覚に大きく貢献するということです。これは新しい発見でした。なぜなら、それ以前は努力感は脳が内部的に生成すると想定されていたからです。この研究の研究者たちは、高い力による疲労性収縮(筋紡錘受容器を感受性低下させる)を用いて被験者の筋紡錘感受性を低下させた後、重りを持ち上げさせました。重りは軽く感じられました——脳の内部的な努力信号が機能しているなら期待されるように重くなるのではなく。また、筋紡錘フィードバックを完全に排除する稀な神経疾患の被験者でも測定しました——疲労後に物体は2倍重く感じられました。健康な被験者では、中枢的な努力推定ではなく、筋紡錘信号(伸展受容器から)が力知覚の支配的な寄与因子であることが示されたのです。
つまり、Luuと同僚たちは局所的な筋肉感覚の現象を研究したわけではありませんが、収縮している筋肉における重さと力の意識的な感覚が、伸展受容器である筋紡錘から来ることを示したのです。
したがって、局所的な筋肉感覚の神経基盤を仮説として立てると、おそらく2つの成分があります:
- II群線維から来る、筋肉がどんな長さにあっても常に存在する緊張性ベースライン
- 錘内筋共活性化によって駆動される、収縮に比例した強化
この感覚の現象学——特に安静時と収縮時の連続性——を分析する中で、私はハムという言葉に行き着きました。なぜなら、まさにハムのようだからです:筋肉が安静時には低いレジスタで存在し、収縮が増すにつれて強まります。同じ現象的な素材が、静かから大きく変化していきます。
Baseworksでは、能動的なタスクにおいてこれらの感覚に注意を向けるとき——具体的には収縮に比例した強化、信号が高まってより読み取りやすくなる部分において——活性化という言葉を使います。しかし「活性化」はその大きな部分だけを捉えています。「ハム」は、安静時のベースラインを含む現象全体を指す言葉として私が使うものです。
これは明示的に仮説です。Luuの論文は力と重さの知覚を扱っており、別個の主観的体験としての局所的な筋肉感覚ではありません。「筋紡錘の再求心性入力が力知覚に寄与する」から「ハムはII群求心性線維が意識的認識に到達したものである」へのステップは論理的ですが、実験的に確認されていません。先に述べた研究者は率直に述べました:3a野を刺激すると安静時の筋肉に局在化できる感覚が生じるという報告は存在しないようです、と。この問いは未解決のままです。
実践においてこれを命名するのに近い別の言葉は「トーン」です。臨床用語の「筋トーン」は、筋肉組織に維持される低レベルの安静時緊張を指します。しかし、広範な動作訓練を積んだ実践者の中には、臨床医が測定するものではなく、自分が感じるものに対してこの言葉を使う人がいます。ダンサーであり動作教育者であるMatan Levkovich(Movement Lab所属)は、まさにこの感覚を「筋トーン」という言葉で表現します——自身のトレーニングでその感覚を受け継いだ言葉として採用したものです。ここでもギターの弦の例えが役立ちます:音楽における「トーン」は、振動できるほど十分に張った弦を指します。錘内筋系が維持するもの——収縮中に筋紡錘を感受性のある状態に保つこと——は、その感覚的等価物のようなものです。
「何も感じていないなら、何もしていないに違いない」
広く見られる運動・トレーニングのマインドセットがあります:強い方が良い、というものです。運動が文化的にどのように促進されているかを見てみると——使われる言葉、何が生産的なセッションと見なされるか、何の感覚が本物の成果のしるしとして提示されているか——強い刺激への根強い強調があります。バーン。ポンプ。努力の感覚。エンドルフィンの放出。長寿への最も効果的な経路としてのウェイトトレーニングへの強調。
この過剰刺激の物語が長期的に多くの人にとってなぜ逆効果かもしれないかについて言えることはたくさんありますが、それはこの記事の範囲外です。ここで焦点を当てたいのは、この物語がさらに筋肉の感覚を隠してしまう要因となり、謎に加わってしまうことです。
筋紡錘の一次求心性線維(Ia群)は、持続的な高い力の収縮中に徐々に感受性が低下します——最大随意収縮の20%での持続的な努力の1分以内に発火率はおよそ半分になり、高い力レベルでは感受性低下がより大きくなります(Macefield et al., 1991)。Luuの実験はこれを直接示しました:高い力による疲労性収縮の後、被験者は持ち上げた重りを軽く感じました(重く感じるのではなく)。筋紡錘信号が鈍くなっていたのです。つまり、運動の物語が有意義なワークアウトと結びつける強度そのものが、活性化の感覚の根底にある可能性がある信号を積極的に低下させるのです。
これを複合させる第二のメカニズムがあります。激しい運動中は、多くの他の信号が皮質の注意を奪い合います:乳酸からの灼熱感(小径のIII群とIV群求心性線維によって運ばれる)、心血管系の興奮、圧倒的な努力感。これらの信号は大きいのです。痛いところをこすると、こすっている間は痛みが軽減されることに気づいたことがあるかもしれません——これは関連した現象で、一つのチャンネルでの強い入力が別のチャンネルでの入力の体験を抑制するものです。これらの信号が大きく走っているとき、微妙で局在化された錘内筋再求心性入力は選択されなくなります——それが存在しないからではなく、注意がそれに向けられる余地がなくなるからです。
そして、最も根本的かもしれない第三のメカニズムがあります。知覚には較正が必要です:より低い強度の信号を見つけるためには、まずより高い強度でそれに注意を向けたことがなければなりません。意識的で読み取りやすいレベルでの筋紡錘のハムを明確に体験したことが一度もない場合——適度な収縮時にそれが顕著で体験に固定できるほど鮮明であるときに注意を向けたことがない場合——安静時にそれを見つけるための知覚的な基準閾値がないのです。安静時の信号は存在します——そしてあなたの脳はその信号を背景で計算に使い続け、「意識的な」位置感覚を届けています——しかし較正された基準がなければ、それは意識的な認識の閾値以下で背景に留まり続けます。これが、ハムが読み取れるほど十分な強度を維持しながら——筋紡錘を感受性低下させたり競合する感覚ノイズを生み出すレベルには到達しない範囲で——練習を保つことが、実際に較正が可能になる条件を作り出す理由の一部です。
全体の連鎖はこのようになります:文化的な物語が高強度へと押し進める→高強度は直接筋紡錘受容器を感受性低下させ、同時に競合する信号で感覚システムを溢れさせる→知覚システムはハムに較正されない→感覚は顕著で注意を向けられた知覚対象にならない→誰もそのための語彙を発展させない→語彙がないから議論がない→議論がないから科学がそれを研究しない→感覚に名前がない。
そして反対の端には——静けさを培うさまざまな「マインドフルネス」の実践や、意図的な収縮なしの非常にゆっくりとした焦点のない動き——があり、ハムの収縮に比例した強化を生み出しません。
そのベースライン信号への意識的なアクセスを発展させるのは別の技法です:収縮に比例した強化を作り出し、それを可能な限り低く意図的に下げていき、今感じたものに対してベースラインの信号を較正できるようにするのです。静けさだけでは、その上昇とベースラインへの帰還を決して作り出しません。活発なダイナミックな動きは、微妙な信号を孤立させるのではなく上書きしてしまう急速な位相変化を生み出す傾向があります。どちらも単独では、安静時に何があるかに注意を向けるために必要な知覚的基準を構築しません。
知覚はその核心においてスキルであり、顕著な細部への注意を必要とするからです:低強度の筋肉感覚への注意が体系的に培われたことがなく、運動に関するあらゆる文化的物語が、本物の仕事のしるしとしてより強い刺激を指し示しているとしたら、この特定の信号が認識された知覚カテゴリになったことがないのは驚くことではありません。
固有受容感覚的意識の有用性とは?
私自身、30歳近くになるまでこれらの感覚に注意を払っていませんでした。そして、Baseworksとその主要な技法のひとつ——あらゆる動きにおいて全筋肉の同時低レベル共収縮を必要とする分散活性化(Distributed Activation)——への関わりが、私の感覚閾値を変え始めました。ほとんど気づかなかったものが、徐々に、そしてかなり実質的に、私の継続的な体験の一部になっていきました。
現在、筋肉から来る感覚と空間における位置感覚は、私の瞬間から瞬間への感覚体験の大きな部分を占めており、純粋に面白く、やりがいが得られるものです。もちろん、時には疑問が湧きます:これは30代に自然に起きることで、練習とは無関係なのかもしれない、と。しかし、祖母も母も、30代になると突然ただ直立して座ることの感覚が魅力的になると教えてくれた記憶がないのです。そして、これらの体験を他の人に伝えようとすると、共通の語彙が欠如していることが多く、共有された体験への言及がないように思えるのです。ソーシャルメディアをスクロールすることからの「ドーパミンヒット」について読むと、本当に?と疑問になるほどです。歩いているときに様々な体の部位の軌道を意識するという、本質的にやりがいが得られる感覚と比較できるのでしょうか?
長い話を短くまとめると:それは非常に心地よく、常に利用可能なのです。
体験の内在的な質を超えて、固有受容感覚的意識は運動学習において直接的な実践的関連性があります。感覚的なアクセスと運動制御の関係は密接に相互接続されているため、単なる知覚的な好奇心以上のものです。
筋肉を感じることができると、その筋肉を制御しやすくなります。さらに、特定の筋肉を単独で制御する能力を訓練すると、その特定の筋肉のための専用フィードバックシステムの発達を伴うことが多いようです。指導の実践において、学生から筋肉の感覚が「沈黙から現れ」「分裂する」という報告を聞きます。これは個別に筋肉を制御できる能力と相関しており、癒合指症の外科的分離後に報告されたものと同様の皮質再マッピングによって説明できるかもしれません(Mogilner et al., 1993)。そのメカニズムは珍しくありませんが、現象学は珍しいのです。誰かが「ここには何も感じなかったのに、今突然感じられるようになった」と言うとき、それは通常、驚きと驚愕を伴います。これをトレーニング成果として追跡するのは容易ではありません——動作範囲や力の出力とは異なり、シンプルな指標や予測可能なタイムラインがありません——しかし指導で聞く報告は一貫しています。
分散活性化(Distributed Activation)——あらゆる動きにおいて、できるだけ多くの筋肉を低強度で共収縮させること——を指導するとき、未経験の実践者は筋肉をどれくらい収縮させれば良いかわからないと報告することが多いです。私の感覚的な観点からは答えは簡単なので、これは興味深いと思います:ただハムに注意を向けて、全筋肉にわたって均一な薄い鞘を作り出せばよいのです。しかし、もちろん、すべての筋肉でそれを知覚していない場合、そして較正されていない場合、これは不可能です。強い感覚なしでも動きを制御することはできますが、固有受容感覚的意識を較正するための訓練は、一般的に個々の筋肉を制御できる能力と相関しています。
実践では、特に分散活性化を学ぶときに、低い固有受容感覚的意識がどのように現れるかの様々なパターンが見られます。共収縮なしにゆるく動く傾向がある人もいれば、まるでストレングストレーニングルーティンの一部として高強度の等尺性ホールドを行っているかのように過活性化する傾向がある人もいます。どちらの場合にも、推測できるのは知覚的なギャップです:正しい活性化レベルの感じ取れる基準がアクセスできない場合、「より多く」または「より少なく」は、実践者が実際に特定して制御できるものに接続されないのです。
固有受容感覚的意識を発達させる方法
ここでの一般的な原則は単純です:受容器があり、その経路が皮質に到達しているならば、その感覚モダリティにおける意識的な弁別能力は練習によって改善できます——対応する感覚に意図的かつ繰り返し注意を向ける行為によって。これは味、色の識別、音にも当てはまります。錘内筋再求心性入力とベースラインの筋紡錘入力にも当てはまらない明白な理由はありません。
ただし、条件が重要です。筋肉神経生理学の研究からわかることを再度まとめると、高強度では関連する受容器が感受性低下し、競合する信号が微妙な錘内筋入力を押しのけます。静止状態では、収縮に比例した成分が生成されません——安静時のハムは存在しますが、意図的な上昇と帰還のコントラストなしに、知覚システムにはそれに較正する機会がありません。瞑想中のボディスキャンは別の知覚層を対象とします——ここで議論している筋骨格系の感覚ではなく内受容感覚に向いています。ボディスキャンは既存のハムのパッチを捉えることができるかもしれませんが、沈黙している筋肉の知覚閾値を引き上げることはありません。
効果があるように思えるのは、それが生み出す感覚に一貫した注意を向けながら、意図的な低〜中程度の強度の活性化です。Baseworksでは、これらの条件が分散活性化(Distributed Activation)とマイクロムーブメント(Micro Movements)を通じて一貫して作り出されます。
なぜこれが機能するかの枠組みは、実践的な仕事自体から生まれました——指導において実際に人々がこれらの感覚へのアクセスを発展させるのに何が役立ったかを何年にもわたって観察することから。私たちはこれを固有受容感覚的意識(Proprioceptive Awareness)と呼びます。なぜなら、科学文献がこの特定の知覚能力に独自の名前を与えていないにもかかわらず、利用可能な最も正確な用語だからです。
そのギャップ——実践で発達できるものと科学が言葉を持つものの間のギャップ——それ自体が謎の一部です。
筋肉活性化感覚アンケート
2022年にこの記事の最初のバージョンを公開したとき、安静時に局所的な筋肉感覚を体験していない人がこれほど多いとは知りませんでした。この知識は、以下のアンケートを通じて蓄積されたデータから得られました。
継続的なデータ収集にご参加いただける方は、以下のフォームからご記入ください。お名前とメールアドレスを入力せず、匿名で完了できます。
参考文献
- Craig AD (2015). How Do You Feel? An Interoceptive Moment with Your Neurobiological Self. Princeton, NJ: Princeton University Press. 10.1515/9781400852727
- Delhaye BP, Long KH, Bensmaia SJ. Neural basis of touch and proprioception in primate cortex. Comprehensive Physiology. 2018 Oct 17;8(4):1575–602.
- Gallagher S. Bodily self-awareness and object perception. Theoria et Historia Scientiarum. 2007;7(1):53.
- Luu BL, Day BL, Cole JD, Fitzpatrick RC. The fusimotor and reafferent origin of the sense of force and weight. J Physiol. 2011;589(13):3135–3147.
- Macefield G, Hagbarth KE, Gorman R, Gandevia SC, Burke D. Decline in spindle support to alpha-motoneurones during sustained voluntary contractions. J Physiol. 1991;440:497–512.
- Mehling WE, Gopisetty V, Daubenmier J, Price CJ, Hecht FM, Stewart A. Body awareness: construct and self-report measures. PLoS One. 2009 May 19;4(5):e5614.
- Mehling WE, Wrubel J, Daubenmier JJ, et al. Body awareness: a phenomenological inquiry into the common ground of mind-body therapies. Philos Ethics Humanit Med. 2009;4:6.
- Mogilner A, Grossman JA, Ribary U, Joliot M, Volkmann J, Rapaport D, Beasley RW, Llinas RR. Somatosensory cortical plasticity in adult humans revealed by magnetoencephalography. Proc Natl Acad Sci. 1993 Apr 15;90(8):3593–7.
- Price CJ, Hooven C. Interoceptive awareness skills for emotion regulation: theory and approach of mindful awareness in body-oriented therapy (MABT). Front Psychol. 2018;9:798.
- Tresilian J. Sensorimotor control and learning: An introduction to the behavioral neuroscience of action. Bloomsbury Publishing; 2012.
- Vallbo AB. Afferent discharge from human muscle spindles in non‐contracting muscles. Steady state impulse frequency as a function of joint angle. Acta Physiol Scand. 1974 Feb;90(2):303–18.
- Vega JA, Cobo J. Structural and biological basis for proprioception. In: Proprioception. IntechOpen; 2021.







